Ters transkriptaz
Biyokimyada bir ters transkriptaz veya RNA'ya bağımlı DNA polimeraz, tek iplikli bir RNA molekülü okuyup (transkripsiyonunu yapıp) tek iplikli DNA üreten bir DNA polimeraz enzimidir. Bu enzim, ayrıca, RNA tek iplikli cDNA şeklinde okunduktan sonra çift iplikli DNA oluşmasında da görev alır. Normal transkripsiyon DNA'dan RNA sentezidir; dolayısıyla ters transkripsiyon bu sürecin tersidir.
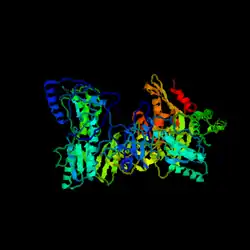 | |
| HIV Ters transkriptazın 3B modeli | |
Ters transkriptaz | |
| Tanımlayıcılar | |
| Sembol | ? |
| Diğer Adlar | Deoksinükleosit-trifosfat: DNA deoksinükleotidiltransferaz (RNA-yönlendirmeli)
|
| Diğer bilgiler | |
| EC numarası | 2.7.7.49 |
Ters transkripsiyon, birbirlerinden bağımsız olarak, University of Wisconsin–Madison'dan Howard Temin ve Massachusetts Teknoloji Enstitüsünden David Baltimore tarafından 1970'te keşfedildi. Bu keşiflerinden dolayı bu iki araştırmacı ve Renato Dulbecco 1975 Nobel Fizyoloji veya Tıp Ödülü'nü paylaştılar.
İyi bilinen ters transkriptazlara aşağıdakiler dahildir:
- İnsan immun yetmezlik virüsü tip I'in HIV-1 ters transkriptazı.
- Moloney fare lösemi virüsü'nün M-MLV ters transkriptazı.
- Kuş miyeloblastosis virüsü'nün AMV ters transkriptazı
- Ökaryotik kromozomların telomerlerini koruyan telomeraz ters transkriptazı
Virüslerdeki fonksiyonu
Ters transkripsiyonlu virüslerde bu enzim ikileşme (replikasyon) sürecinde kullanılır. Ters transkripsiyonlu RNA virüslerinde bu enzim RNA genomlarının DNA'ya dönüştürülmesinde kullanılır, bu DNA sonra konak organizmanın genomu ile bütünleşir ve onunla beraber ikilenir. Ters transkripsiyonlu DNA virüsleri, hepadnavirüsler gibi, ters transkripsiyon sayesinde DNA ipliklerinin üretimi ve birleştirilmesinde RNA'yı bir kalıp olarak kullanır. AIDS hastalığının virüsü HIV, bu enzimi kullanarak insanları enfekte eder. Ters transkriptaz olmazsa viral genom konak hücrenin içine dahil olamaz, bu da onun çoğalamaması demektir.
Ters transkripsiyon süreci
Ters transkripsiyon bir RNA kalıptan tek zincirli bir DNA sentezlenmesidir, buna RNA yönlendirmeli DNA sentezi (RNA-directed DNA syntesis)) de denir. Çoğu ters transkriptaz enziminde bu etkinliğe ek olarak DNA yönlendirmeli DNA sentez yeteneği de olur. Retrovirüslerdeki ters transkriptazlarda ayrıca bir RNaz H etkinliği vardır, bu bir DNA-RNA hibritindeki RNA zincirinin sindirilmesidir.
DNA'ya bağımlı DNA polimeraz etkinliği olmayan, ters transkriptazlı virüslerde iki iplikli DNA'nın oluşturulması konak hücre tarafından kodlanan DNA polimeraz δ tarafından mümkün olur. Bu enzim, viral DNA-RNA kompleksindeki RNA'yı bir primere benzetip iki iplikli bir DNA sentezler, bu süreç sırasında yeni sentezlenen DNA orijinal RNA kalıbı yerinden çıkarır.
Ters transkripsiyon çok hataya eğilimlidir ve bu süreç zarfında mutasyonlar meydana gelebilir. Bu mutasyonlar antiviral ilaç direncine neden olabilir.
Telomeraz
Telomerazlar kromozomların ucuna kısa DNA parçaları ekleyerek (omurgalılarda ""TTAGGG" dizileri) kromozomların her ikileşmesini takiben meydana gelen kısalmayı düzeltirler. Telomeraz enzimleri kendi yapıları içinde bir RNA kalıp molekül taşırlar. Bu RNA kalıp 3'-CAAUCCCAAUC-5' dizisine sahiptir. Telomeraz kromozomun ucuna bağlanıp bu RNA kalıbı kromozomun ucundaki en son TTAGGG ile hizalar, sonra kalıp ile komplementer bir DNA zinciri sentezler. Bu işlemin tekrar tekrar yapılması ile kromozom uzar.
Sınıf VI virüslerdeki süreç
Sınıf VI virüsler retrovirüs olarak adlandırılırlar, bunlar DNA araürünlü RNA ters transkriptaz virüslerdir. Genomları pozitif anlamlı, 5' başlıklı ve 3' poliadenile kuyruklu iki adet tek iplikli RNA molekülünden oluşur. Retrovirüslere örnek olarak İnsan Bağışıklık Yetmezlik Virüsü (HIV olarak adlandırılır) ve İnsan T-Limfotropik Virüs (HTLV) gösterilebilir. Ters transkripsiyon sitozolda gerçekleşir.[1] Retroviral ters trankriptazların çalışma mekanizması telomerazlardn çok daha karmaşıktır. Çift iplikli DNA'nın oluşumu aşağıdaki adımlarla olur (sağdaki şekle bakınız):
- Spesifik bir hücresel tRNA bir primer olarak etkir ve virüs genomunun komplementer bir kısmı ile hibritleşir. Buraya primer bağlanma yeri (PBY) olarak adlandırılır.
- Primerin sağladığı -OH grubuna nükleotitlerin eklenmesi ile komplementer bir DNA zinciri (cDNA) sentezlenir. Viral RNA'nın 5' ucundaki U5 (kodlamayan bölge) ve R bölgesine (RNA molekülünün her iki ucundaki doğrudan tekrarlı bölge) DNA olarak kopyalandıktan sonra sentez sona erer.
- Ters transkriptaz enziminin RNaz H olarak adlandırılan bölgesi, DNA-RNA kompleksindeki RNA'yı sindirir, böylece viral RNA'nın 5' ucundaki U5 ve R bölgeleri ortadan kalkar.
- Ucunda yeni sentezlenmiş U5 ve R bölgelerinin DNA'sı olan primer, viral RNA'dan ayrışır ve kendisine komplementer olan, viral genomun 3' ucundaki R bölgesi ile hibritleşir.
- Komplementer DNA'nın uzatılır.
- Viral RNA'nın büyük kısmı (PP bölgesi hariç) RNaz H tarafından sindirilir.
- İkinci DNA ipliğin sentezi viral RNA'nın arta kalan kısmından (PP bölgesinden) başlatılır.
- Birinci DNA ipliğin primeri olan tRNA yıkıma uğrar.
- İki DNA ipliği ayrışır, ikinci ipliğin PBY'si birinci iplikteki komplementer PBY ile hibritleşir. Viral RNA'nın arta kalan son kısmı (PP bölgesi) de yıkıma uğrar.
- Her iki iplik daha da uzatılarak çift iplikli DNA oluşur.
- Sentezi tamamlanan viral DNA, entegraz enzimi aracılığıyla konak genom ile bütünleşirler.
İki iplikli DNA'nın oluşması iplik aktarımı ile olur, bu süreçte RNA'ya-bağımlı DNA sentezinin ürünü olan kısa DNA, genomun öbür ucundaki kalıp bölgelere taşınır, bu bölgelere sonradan ulaşan ters transkriptaz kendi DNA'ya-bağımlı DNA polimeraz aktivitesi için bunları işlemler.[2]
Retroviral RNA'da primerin bağlandığı yere primer bağlanma yeri (PBY) denir. PBY'nin 5' yönündeki bölge U5 olarak adlandırılır, PBY'nin 3' tarafındaki kısmı ise lider olarak adlandırılır. tRNA çift ipliklikçikli kısmından 14 ila 22 nükleotit kadar açılarak PBY'de viral RNA ile hibritleşir. PBY'nin viral RNA'nın 5' ucunda yer alması diğer virüslere göre sıradışıdır çünkü ters transkriptaz DNA'yı sentezlerken, primerin 3' ucundan başlayarak, 5'-3' doğrultusunda ilerler ve viral genomun sadece ufak bir kısmını kopyalamış olur. Dolayısıyla, primer ve ters transkriptaz, RNA'nın 3' ucuna taşınmalıdır. Bu yer değiştirmenin gerçekleşmesi için, çeşitli enzimler (DNA polimeraz, RNaz H gibi) ve yukarıda belirtilen çok adımlı bir süreç gerekir.[3]
HIV ters transkriptazının ayrıca bir ribonükleaz aktivitesi vardır, bu aktivite cDNA sentezi sırasında kalıp olarak kullanılmış olan viral RNA'yı yıkıma uğratır, bir de DNA'ya bağımlı DNA polimeraz etkinliği vardır, bunun sayesinde viral RNA'nın ters transkripsiyonu ile elde edilen anlamlı cDNA ipliğini antianlamlı DNA olarak kopyalayarak çift iplikli bir viral DNA (vDNA) araürünü oluşturur.[4]
Ökaryotlarda
Ökaryotik genomlarda retrotranspozon olarak adlandırılan, kendini ikileyen bölgeler vardır. Bunlar bir ters transkriptaz kullanarak, bir RNA araürün aracılığıyla kendilerini genomun bir yerinden başka bir yerine taşırlar. Hem hayvan hem de bitki genomlarından çok sayıda bulunurlar. Çoğu ökaryotta (insan da dahil) bulunan bir diğer ters transkriptaz telomerazdır, bu enzim kendi RNA kalıbını taşımaktadır; telomerazın RNA'sı, DNA ikileşmesi için kalıp olarak kullanılır.[5].
Prokaryotlarda
Bazı bakterilerde Retron msr RNA olarak adlandırılan, ters transkriptaz kodlayan ayrık diziler bulunur, bunlar çok kopyalı tek iplikli DNA (İngilizce multicopy single-stranded DNA, msDNA) sentezi için kullanılırlar. DNA sentezinin başlaması için bir primer gerekir. Bakterilerde bu primer ikileşme sırasında sentezlenir.[6]
Yapı
Ters transkriptaz enziminde RNA'ya bağımlı DNA polimeraz ve DNA'ya bağımlı DNA polimeraz bölgeleri, bunlar birlikte çalışarak transkripsiyon yaparlar. Transkripsiyon işlevine ek olarak, retrovirüslerin ters transkriptazlarında RNaz H protein ailesine ait bir bölge de bulunur, bu da onların ikileşmesi için hayatî önem taşır.
İkileşme sadakatı
Bir retrovirüsün hayat döngüsü sırasında üç farklı ikileşme (replikasyon) sistemi vardır. Birincisi, ters transkriptaz viral RNA'dan başlayarak, peşpeşe iki DNA ipliği sentezleyerek viral DNA'yı oluşturur. İkinci replikasyon sürecinde konak DNA polimeraz entegre olmuş viral DNA'yı ikiler. Son olarak, RNA polimeraz II, proviral DNA'yı RNA olarak yazar (transkripsiyonunu yapar), bu RNA da viryonların içinde paketlenir. Bu ikileşme adımlarının her birinde mutasyonlar meydana gelebilir.[7]
Ters transkriptaz, RNA'yı DNA olarak yazarken yüksek bir hata oranı gösterir çünkü, DNA polimerazdan farklı olarak, bu enzimin prova okuma yeteneği yoktur. Hata oranı örneğin AMV ters transkriptazının için 1/17.000 baz, M-MLV ters transkriptazı için de 1/30.000 bazdır.[8]
Uygulamalar
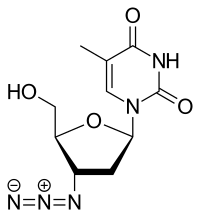
Antiviral ilaçlar
HIV ters transkriptaz kullanarak genetik malzemesini kopyaladığı ve yeni virüsler ürettiği için, bu süreci bozacak ve böylece büyümesini engelleyecek spesifik ilaçlar tasarlanmıştır. Toplu olarak bu ilaçlar ters transkriptaz inhibitörleri olarak adlandırılır. Bunların arasında nükleosit veya nükleotit analogları olan zidovudine (tescilli marka adı Retrovir), lamivudin (Epivir) ve tenofovir (Viread) ve nükleosit olmayan inhibitörler, nevirapine (Viramune) gibi, sayılabilir.
Moleküler biyoloji
Ters transkriptaz moleküler biyoloji laboratuvarlarında yaygın kullanılan bir enzimdir, özellikle ters trasnkriptaz polimeraz zincir tepkimesi (İng. reverse transcription polymerase chain reaction veya RT-PCR) diye adlandırılan ve polimeraz zincir tepkimesinin RNA'ya uygulandığı bir yöntemde kullanılır. Klasik polimeraz zincir tepkimesi (İng. polymerase chain reaction veya PCR) sadece DNA zincirlerine uygulanır ama, ters transkriptaz aracılığıyla RNA'dan DNA sentezlenirse PCR kullanımına uyarlanmış olur. Ters transkriptaz ayrıca mRNA'dan cDNA kitaplıkları üretilmesine olanak verir. Ters transkriptazın ticarî üretimi moleküler biyolojide yeni bilgi üretimine büyük katkıda bulumuştur, çünkü, bu enzim (başka enzimlerle birlikte) araştırmacıların RNA dizilerinin DNA olarak klonlanıp dizilemesini ve karakterize edilmelerini mümkün kılmıştır.
Tarihçe
Howard Temin ve David Baltimore birbirinden bağımsız 1970 yılında ters transkriptazı ilk olarak keşfettiler. Bunun için iki bilim adamı Renato Dulbecco ile beraber Nobel Fizyoloji veya Tıp Ödülünü aldılar. Keşfettikleri enzimle aynı zamanda proteinlerin DNA'dan RNA oluşmasından sentez edilmediğini ispatladılar. O zamanlar mikrobiyologlar arasında bu görüş çok yaygındı. Bu görüşün tersini, yani RNA'dan DNA oluşmasından proteinlerin sentez edilmesini keşfettikleri için bu enzime "ters transkriptaz" adını verdiler.[9]
Ayrıca bakınız
- cDNA kitaplığı
- DNA polimeraz
- msDNA
- Ters transkripsiyonlu virüsler
- RNA polimeraz
- Telomeraz
- Retrotranspozon belirteç
Dış bağlantılar
- Ters transkriptaz ve üç ters inhibitörünün animasyonları
- Protein Data Bank'ta Ayın molekülü (Eylül 2002)
- BRENDA veritabanı girdisi - bir protein veritabanındaki ayrıntılı bilgiler
- EC 2.7.7.49
- HIV ikileşmesi (Kasım 2008). Boehringer Ingelheim tarafından hazırlanmış video.
Kaynakça
- Bio-Medicine.org - Retrovirus erişimtarihi=?7 Şubat 2009
- Telesnitsky, A., Goff, S.P. (1993). "Strong-stop strand transfer during reverse transcription". Skalka, M. A., Goff, S.P (Ed.). Reverse transcriptase (1 bas.). New York: Cold Spring Harbor. s. 49. ISBN 0-87969-382-7.
- Weiss, R., N. Teich, H. Varmus, and J. Coffin (1985). "RNA tumor viruses". Jonathan, L., Ashock, A., David, C. (Ed.). Molecular biology of tumor viruses (2 bas.). New York: Cold Spring Harbor.
- Doc Kaiser's Microbiology Home Page > IV. VIRUSES > F. ANIMAL VIRUS LIFE CYCLES > 3. The Life Cycle of HIV 26 Temmuz 2010 tarihinde Wayback Machine sitesinde arşivlendi. Community College of Baltimore County. Updated: Jan., 2008
- Lodish, et al, Molecular Cell Biology (2004), 5th edn, W. H. Freeman and Company, New York, ISBN 0-7167-4366-3
- Hurwitz, J. and J. Leis (1972). "Directing influence of DNA in the reaction". RNA-dependent DNA polymerase activity of RNA tumor viruses. ss. 116-129.
- Bbenek, K., Kunkel, A. T (1993). "The fidelity of retroviral reverse transcriptases". Skalka, M. A., Goff, P. S. (Ed.). Reverse transcriptase. New York: Cold Spring Harbor Laboratory Press. ss. p. 85. ISBN 0-87969-382-7.
- "Promega kit instruction manual (1999)" (PDF). 21 Kasım 2006 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 11 Ağustos 2010.
- John M. Coffin, Stephen H. Hughes, Harold E. Varmus: The Place of Retroviruses in Biology., Retroviruses. Cold Spring Harbor Laboratory Press, 1997, ISBN 0-87969-571-4.