Krosover
Crosover veya krossing over ya da parça değişimi mayoz bölünmenin profaz I evresinde görülen, çift halde bulunan kromozomların yaptığı parça değişimine verilen addır. Bunun sonucunda genetik rekombinasyon meydana gelir, yani farklı kromozomlarda bulunan genlerin alelleri birbiriyle yer değiştirir.
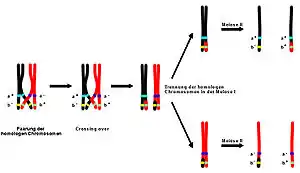
Krosover, genelde sinapsis denen bir süreç zarfında olur. Sinapsis, özellikle pakiten evresinde sinaptonemal kompleksin oluşumundan önce başlar ve profaz I'in sonuna kadar tamamlanmaz. Krosover genellikle, eşleşmiş kromozomların eşleşme bölgelerinin kırılıp sonra tekrar diğer kromozomla birleşmesi sonucu meydana gelir. Bu genleri içeren parça değişiminin sonucu genler iki kromozom arasında yer değiştirir, buna genetik rekombinasyon denir. Bu süreç sırasında birbirine benzer iki DNA parçası aynı noktadan kırılır, bunların her biri karşı zincirin öbür parçasıyla birleşerek kırıklarını onarırlar.
Tarih
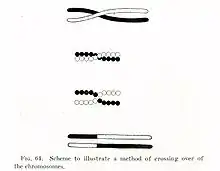
Belçikalı Frans Alfons Janssens bölünen hücrelerin kromozomlarında "kiyazma" diye adlandırdığı yapılar gözlemlemiştir. Kiyazma, krosoverin içinde yer gerçekleştiği fiziksel yapıdır. Thomas Hunt Morgan, kendi Drosophila deney sonuçlarını açıklamakta Janssens'in gözleminin önemini hemen kavramış, ilk defa krosover mekanizmasını teorik olarak tarif etmiştir. Krosoverin fiziksel temellerini ise Harriet Creighton ve Barbara McClintock 1931'de göstermiştir.[1]
Morgan bu sürece İngilizce 'karşıya geçme' anlamında "cross-over" adını vemiştir, bu 'krosover' veya 'krossing over' olarak Türkçeleşmiştir.
Kimyası
Mayozik rekombinasyon, Spo11 proteinin DNA çift zincirini kırmasıyla başlar.[2] Bir ya da daha fazla eksonükleaz, kırılmış çift zincirin 5' uçlarını sindirerek, DNA'da 4' uçlu uzantılar olmasını sağlar. Mayoza özgül rekombinaz Dmc1 ve genel rekombinaz Rad51 tek zincirli DNA'nın üstünü kaplayıp filamentler oluştururlar.[3]
Rekombinaz enzimleri DNA'nın tek zincirli ucunun karşı taraftaki kromatitin arasına girmesini (istila etmesini) katalizler. İstilacı DNA'nın 3' ucu DNA sentezini başlatır ve tamamlaycı iplikçiğin kenara iter. Kenara itilen tamamlayıcı iplikçik baştaki iki zincirli kırığın öbür tarafındaki tek iplikçikli DNA ile eşleşir. Ortaya çıkan yapıya karşılıklı iplikçik değiş tokuşu denir (İng. cross-strand exchange) ve "Holliday kavşağı" olarak isimlendirilir. Birbirine temas eden ve yakında krosover yapacak iki kromatitin temas noktasına kiyazma denir. Holliday kavşağı, her iki tarafından rekombinazlarla çekilen dörtlü bir yapıdır.
Krosoverin sonucu
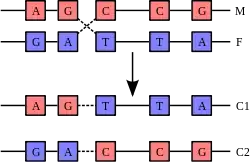
Krosover sırasında homolog kromozomların biri anneden diğeri babadan gelmiştir. Bu şekilde iki farklı bireyin kromozomlarındaki genetik bilgi birbirine karışarak yeni kombinasyonlar oluşturur. Bu parça değişimi olmasa, canlının kromozomundaki genetik bilgi mutasyonlar haricinde değişmezdi.
Çoğu ökaryot, hücrelerinde genlerinin iki kopyasını taşır ve bunların her birine allel denir. Her ebeveyn yavrusuna birer allel aktarır. Metafaz sırasında dizilen kromatit çiftlerinin her birinden birer tane alınması sonucunda, bir gamet, bütün kromozomlardaki allelerin haploit bir bileşimini ebeveyn hücreden kalıt alır. Rekombinasyon olmazsa, aynı kromozom üzerinde bulunan tüm alleler beraber aktarılır. Mayotik rekombinasyon, kardeş kromatitler üzerinde bir gene ait iki farklı allelin değişimi sonucunda alellerin daha serbestçe karışmasını sağlar.
Rekombinasyon olması halinde, bir yavrudaki allel dağılımının bir diğerindeki ile aynı olma olasılığı aynıdır. Buna Mendel genetiğinde "bağımsız tertiplenme" kanunu denir. Farklı kromozomlarda bulunan genlerin allellerinin dağılması ile, aynı kromozomda olan ve aralarında her zaman rekombinasyon olan genlerin allellerinin dağılması aynı davranışı gösterir. Ancak bu kanunun bir istisnası vardır.
Bir kromozom üzerinde iki gen arasında bir rekombinasyon olma olasılığı her gen çifti için aynı değildir. İki gen birbirine çok yakın ise bir rekombinasyon olayının bu ikisini birbirinden ayırma olasılığı, iki genin birbirinden uzakta olma durumuna kıyasla daha düşüktür. "Genetik mesafe", rekombinasyon sıklığının yeterince çok sayıda soy ağacına bakılarak elde edilen ortalama değeridir. Genetik bağlantı (İng. genetic linkage), aynı kromozomdaki genlerin konumlarından dolayı beraber kalıt olma eğilimini betimler. Bağlantı dengesizliği (İng. Linkage disequilibrium) bazı gen veya gen belirteci (İng. gene markers) kombinasyonlarının aralarındaki mesafeye göre olması gerekenden daha sız veya daha ender olma durumlarıyla ilgilidir. Belli bir hastalığa nedeni olan gen aranırken bu kavramdan yararlanılır. Bunun için, bir DNA dizisinin belli bir hastalıkla beraber rastlanmasına bakılır; eğer ikisi arasında yüksek bir korelasyon bulunursa aranan genin bu DNA dizisinin yakınında olduğunda hüküm verilir.
Problem
Krosover genelde eşleşen kromozomların benzeşen (homolog) bölgeleri arasında gerçekleşse de DNA dizi benzerlikleri yüzünden bazen yanlış noktalarda hizalanma olabilir. Bu durumda dengesiz rekombinasyon meydana gelir. Normal rekombinasyona kıyasla dengesiz rekombinasyon oldukça ender olur, ama dengesiz rekombinasyonlar içeren bir gametten bir zigot oluşursa, ciddi sorunlar meydana gelebilir. Bunun sonucunda bir kromozomdaki genlerin düplikasyonu ve bir diğerindekilerin delesyonu, veya bir kromozom parçasının başka bir kromozoma translokasyonu veya evrilmesi (inversiyonu) olabilir.
Kaynakça
- Creighton H, McClintock B (1931). "A Correlation of Cytological and Genetical Crossing-Over in Zea Mays". Proc Natl Acad Sci U S A. 17 (8). ss. 492-7. PMID 16587654.
- Keeney S, Giroux CN, and Kleckner N (1997). "Meiosis-specific DNA double-stranded breaks are catalyzed by Spo11, a member of a widely conserved protein family." Cell 88(3):375-384. PMID 9039264 DOI:10.1016/S0092-8674(00)81876-0
- Sauvageau S, Stasiak AZ, Banville I, Ploquin M, Stasiak A, and Masson JY (2005). "Fission yeast rad51 and dmc1, two efficient DNA recombinases forming helical nucleoprotein filaments." Mol Cell Biol 25(11):4377-4387. PMID 15899844 DOI:10.1128/MCB.25.11.4377-4387.2005
Ayrıca bakınız
![]() Wikimedia Commons'ta Krosover ile ilgili çoklu ortam kategorisi bulunur.
Wikimedia Commons'ta Krosover ile ilgili çoklu ortam kategorisi bulunur.