Meselson-Stahl deneyi
Meselson-Stahl deneyi, DNA ikileşmesinin yarı korumalı olduğu hipotezini destekleyen, Matthew Meselson ve Franklin Stahl tarafından yapılmış bir deneydi. Yarı korumalı ikileşmenin (replikasyonun) anlamı, DNA sarmalının iki ipliği ikileşince, meydana gelen her bir çift iplikli DNA sarmalındaki ipliklerden birinin orijinal sarmaldan geldiği, öbürünün ise yeni sentezlenmiş olduğudur. 1958'te yayımlanan bu deneyin sonuçları moleküler biyolojinin en önemli deneyleri arasında yer alır.
Hipotezler
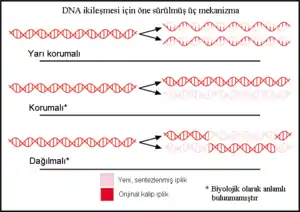
DNA'nın ikileşme yöntemi hakkında üç hipotez öne sürülmüştü.
Yarı korumalı hipoteze göre, ikileşme sırasında DNA'nın iki iplikçiği ayrılır, her iplikçik yeni bir iplikçiğin sentezi için kalıp görevi görür. Bu hipotez Watson ve Crick tarafından öne sürülmüştü.[1].
Korumalı hipoteze göre, DNA molekülün tümü tamamen yeni bir molekülün sentezlenmesi için kalıp görevi görür. Bu modele göre, DNA'ya bağlı olan histon proteinleri onun şeklini bozarak her iki iplikteki bazları hidrojen bağlanması için ortaya çıkarırlar.[2].
Dağılmalı hipotezin bir örneği, Max Delbrück tarafından önerilen bir modelde görülebilir. Bu modele göre birbirine sarılı durumda olan iki iplikçiğin açılması sorununun çözümü için DNA omurgası her 10 nükleotitte bir kesilmesini sağlayacak bir mekanizma sonucu molekülün gevşeyebildiğini savunur. Eski iplikçiğin parçaları yeni sentezlenmekte olan yeni iplikçiğin ucuna eklenir. Eski molekülden kısa parçalar kah bir iplikçikten kah öbür iplikçikten gelerek yeni sentezlenen molekülü oluştururlar.[3].
Bu üç modelin her biri "eski" DNA molekülünün yeni oluşan moleküller içindeki dağılımı hakkında farklı öndeyilerde bulunmaktadır. Korunmalı hipotezde, ikileşme sonucuda meydan gelen moleküllerden biri tamamen korunmuş olan "eski" molekülde oluşur, öbürü ise yeni sentezlenmiş DNA'dır. Yarı korunmalı hipoteze göre ikileşmenin ardından her yeni molekülde bir eski ve bir yeni iplik bulunacaktır. Dağılmalı modele göre ise yeni molekülün her bir ipliği eski ve yeni DNA'nın bir karışımıdır.[4]
Deneysel yöntem ve sonuçlar
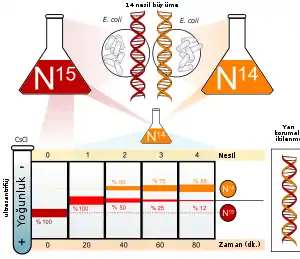
Azot, DNA'yı oluşturan başlıca elementlerden biridir. 14N , azotun en yaygın izotopudur ama ondan daha ağır olan (ve radyoaktif olmayan) 15N izotopunun içeren DNA'da kimyasal olarak aynı özelliklere sahiptir.
E. coli bakterisi 15N içeren bir ortamda birkaç nesil boyunca büyütülür. Bu hücrelerden DNA elde edilip bir tuz yoğunluk gradyanında santrifüjlenince, DNA molekülleri tuz çözeltisi içinde ilerleyip çözeltinin kendileriyle aynı yoğunlukta olduğu noktada dururlar. 15N'li ortamında büyümüş hücrelerden elde edilen DNA'nın yoğunluğu, normal 14N'li ortamda büyümüş hücrelerdekinden daha yüksektir. Bunun ardından, DNA'larında sadece 15N bulunan E. coli hücreleri 14N'lü bir ortama aktarıldılar ve orada büyümelerini sürdürdüler. Hücrelerin çoğalmasını izlemek için hücre süspansiyonunun optik yoğunluğu ölçüldü.
DNA bu hücrelerden düzenli aralıklarla özütlendi (elde edildi) ve saf 14N DNA ve 15N DNA ile karşılaştırıldı. Bir hücre bölünmesinin ardından DNA'nın yoğunluğu ara bir bir değere sahipti. Korumalı ikileşme modeline göre eşit miktarda yüksek ve alçak yoğunluklu DNA meydana gelmesi gerektiğine göre, bu hipotez çürütülmüş oldu. Fakat bu sonuç hem yarı kormalı hem de dağıtmalı ikileşme ile tutarlı idi. Yarı korumalı ikileşmeye göre iki iplikli DNA'nın bir ipliği 15N'li, öbürü 14N'li olmalıdır, dağılmalı ikileşme modeline göre ise her iki iplikte 15N ve 14N karışık durumda olmalıdır, yani her iplik ara değerde bir yoğunluğa sahip olmalıdır.
Araştırmacılar hücreler bölündükçe numuneler almaya devam ettiler. İki bölünmeden sonra elde edilen DNA'nın eşit oranda iki farklı yoğunluk değerine sahip olduğu bulundu. Bunlardan biri, sadece bir kere bölünen hücrelerden elde edilen DNA'daki gibi bir ara yoğunluk değerine sahipti, öbürü ise tamamen 14N'lü ortamda büyüyen hücrelerden elde edilen DNA'nın yoğunluk değerine sahipti. Bu sonuç dağılmalı ikileşme hipotezi ile uyumsuzdu, çünkü bu hipoteze göre tüm DNA moleküllerinin aynı yoğunluğa sahip olması gerekirdi, bu yoğunluk değeri bir kere bölünmüş hücrelerdeki yoğunluk diğerinden daha düşük olmalıydı ama sırf 14N'lü ortamda büyümüş bakterilerin DNA'sının yoğunluğundan yüksek olmalıydı, çünkü orijinal 15N'li DNA, yeni iplikler arasında paylaşılmış olmalıydı. Sonuç yarı korumalı ikileşme hipotezi ile uyumlu idi.[5].
Kaynakça
- WATSON JD, CRICK FH (1953). "The structure of DNA". Cold Spring Harb. Symp. Quant. Biol. Cilt 18. ss. 123-31. PMID 13168976.
- Bloch DP (Aralık 1955). "A POSSIBLE MECHANISM FOR THE REPLICATION OF THE HELICAL STRUCTURE OF DESOXYRIBONUCLEIC ACID". Proc. Natl. Acad. Sci. U.S.A. 41 (12). ss. 1058-64. PMC 528197 $2. PMID 16589796.
- Delbrück M (Eylül 1954). "ON THE REPLICATION OF DESOXYRIBONUCLEIC ACID (DNA)". Proc. Natl. Acad. Sci. U.S.A. 40 (9). ss. 783-8. PMC 534166 $2. PMID 16589559.
- Delbrück, Max (1957). "On the mechanism of DNA replication". William D. McElroy, Bentley Glass (Ed.). A Symposium on the Chemical Basis of Heredity. Johns Hopkins Press. ss. 699-736.
- Meselson, M. and Stahl, F.W. (1958). "The Replication of DNA in Escherichia coli". PNAS. Cilt 44. ss. 671-82. doi:10.1073/pnas.44.7.671. PMID 16590258.
Dış bağlantılar
- DNA From The Beginning Deneyi açıklayan bir animasyon.
- The Meselson–Stahl Experiment Bir diğer yararlı animasyon.