Gen duplikasyonu
Gen duplikasyonu (veya kromozom duplikasyonu veya gen amplifikasyonu veya gen ikilenmesi), içinde bir gen bulunan bir DNA bölgesinin herhangi şekilde ikilenmesidir; homolog rekombinasyon sırasında bir hata sonucu, retrotranspozisyon olayı veya tüm bir kromozomun ikilenmesi sonucu meydana gelebilir.[1] Genin kopyası selektif baskıdan yoksun olduğu için, ondaki mutasyonların organizma üzerinde zararlı etkisi olmaz. Dolayısıyla, organizmanın nesilleri boyunca, işlevsel tek kopyalı bir gene kıyasla daha hızlı mutasyona uğrar.
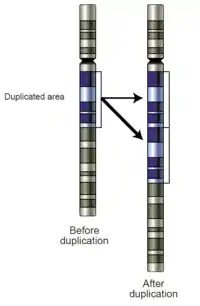
İkilenmenin meydana gelme mekanizmalarından biri hatalı rekombinasyondur. Bu durumda ikilenme ve silinme (delesyonun) beraber meydana gelir. Mayoz bölünme sırasında homolog kromozomların hatalı hizalanması sonucu eşitsiz krosover olarak adlandırılan bir olay olur. Bu rekombinasyonun sonucu, değişim yerindeki kromozomlardan birinde bir ikilenme, öbüründe ise bir delesyon oluşur. [2]
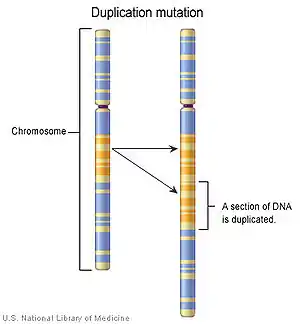
Duplikasyon, bir kromozomun bir parçasının o kromozom üzerinde iki veya daha fazla sayıda tekrarla görülmesi şeklindeki kromozom anomalisidir. Yani kromozomun bir kısmının kendi kendini eşlemesi olarak da tanımlanabilir.
Duplikasyonlu bir parça sentromerli serbest bir parça veya tamamlayıcı bir kromozom parçası olabilir. Eğer sentromere sahip, yani sentrik bir kromozom parçası ise, bu parça küçük, ekstra bir kromozom olarak kabul edilir.
Bir kromozom parça değişimi sırasında karşısındakine belirli genleri vermez, sadece alırsa o gen bakımından diploid olur. Bu, çoğunlukla düzenli işlemeyen bir krossingoverde meydana gelir. Normal olarak mayozun ilk evrelerinde eş genler sinapsis yapar. Ayrılırken normal bir bölünme olmazsa, kromatidlerden biri, o gen bakımından diploid olur, diğeri ise o genlerden yoksun kalır.
Homolog kromozomlar ilk mayoz sırasında sinapsis sonrası ayrılırken farlık noktalarından çaprazlandıklarında koma ve değişik düzeylerde kaynaşma olursa iki tip kromozom ortaya çıkar. örneğin ABCDE kromozomunun BC lokusları ile homoloğunun DE lokusları arasında bir kiazma noktası oluşursa, kırılma ve parça değişimi sonunda biri ABE, diğeri de ABCDCDE genlerini içeren kromozomlar oluşur. Buradaki parça değişimi nokta nokta karşılıklı gelme şeklinde olan krossingoverden farklıdır. Duplikasyon yani iki tane normal homoloğu ile karşılıklı geldiğinde bu fazla parça kısmı dışarıya doğru bir halka yapısı gösterir. Dolayısıyla Profaz I sırasında gözlenen ilmek veya halka yapıları delesyon veya duplikasyon olaylarına işaret eder.
Birbirini takip eden ikilenmelere en iyi örnek Drosophila sineğinde "Bar Duplikasyonu" diye bilinen durumdur. Drosophila'larda bulunan Bar mutant geni petek gözün küçülmesine sebep olmaktadır. de Marinis adlı bir araştırmacı Drosophila X kromozomununda bir parçanın iki defa bulunduğunu göstermiştir. 16 A diye adlandırılan bu parça, düzensiz bir krossingover sonucu bir kromatidde tek diğerinde ise üç tane bulunabilmektedir. Özel markalama yöntemleri ile bu durum gösterilebilmiştir.
Her bir çekirdekte 16 A segmentinin sayısı aynı olsa da 16 A'nın kromzomlardaki yerleşim durumları da önemlidir. Örneğin bir çekirdek içinde bir kromozomda üç, onun homoloğunda bir tane 16 A segmenti taşıyan hayvanlardaki petek gözü oluşturan kafes sayısı, çekirdeklerin de her birinde ikişer tane 16 A segmentli kromozom çifti taşıyan sineklerin petek göz kafes sayılarından ile sembolize edildiğinde, normal durumlarda ABCDEFGH, bar özelliğinde ABCDEFGBCDEFGH, ultrabar özelliğinde ise, ABCDEFGEBCDEFGBCDEFGH gen içeriği vardır.
Genellikle ikilenmeler şeklindeki gen sayısı atışları, parça delesyonlarına nazaran daha az zaralıdır. Çünkü hücrede orijinal gen kombinasyonunu bozmadan DNA ilavesi mutasyonun oluşmasını sağlar.
Duplikasyon, evrim bakımından önemlidir. Çünkü hücreye ek DNA girmesini ve böylece orijinal gen kompleksini bozmadan mutasyonların meydana gelmesini sağlar. Genellikle zararlı bir etkisi yoktur fakat bazen gen dengesini bozduğu için zararlı sonuçlar doğurabilir. Örneğin, Drosophila'da gözlerin küçük kalmasını sağlayan "Bar Duplikasyonu" buna tipik örnektir. Bitkilerde devleşme meydana getirebilir. Genellikle kararlı olmayan değişimlerdir, zamanla normal durumuna dönüşebilir.
Bloom ve Rett sendromu kromozomal duplikasyon sonucu meydana gelen genetik hastalıklardandır.
Evrimsel bir olay olarak gen duplikasyonu
Gen duplikasyonunin evrimde önemli bir rol oynadığına en az yüz yıldır inanılmaktadır.[3] Susumu Ohno klasik eseri Gen Duplikasyonu ile Evrim[4] ile bu teorinin en meşhur geliştiricilerindendir. Ohno, evrensel ortak atanın]] ortaya çıkmasından beri en önemli evrimsel kuvvetin gen duplikasyonu olduğunu savunmuştur.[5] Büyük genom ikilenme olayları ender değildir. Ekmek mayasının genomunun yaklaşık 100 milyon yıl önce ikilendiğine inanılmaktadır.[6] Bitkiler en sık genom ikilemesi yapan canlılardır. Örneğin buğday heksaploittir (bir poliploid çeşidi), yani genomunun altı kopyasına sahiptir.
Bir gen ikilenme olayı ile meydana gelen iki gen paralog olarak adlandırılır ve genelde farklı yapı ve işlevi olan proteinler kodlarlar. Buna karşın ortolog genler farklı canlı türlerinde bulunan benzer işlevli proteinleri kodlar ama bunlar bir türleşme olayı sonucu meydana gelmişlerdir. (Bakınız Genetikte dizi homolojisi.)
Paralog ve ortologların ayırdedilmesi biyolojik araştırmalarda önemli ama zordur. Bir insan geninin homoloğu başka bir canlı türde bulunabilirse, o canlıda deney yaparak insan geninin fonksiyonu hakkında bilgi edinilebilir, ama ancak bu homoloji ortolog ise. Eğer homolog genler paralog iseler ve bir gen ikilenme olayı sonucu meydana geldilerse, işlevleri muhtemelen birbirlerinden çok farklıdır.
Paralog parçalar, birbirlerine %90 benzerlik gösteren tekrar dizileri olabilir. Böyle durumlarda bu paralog bölgeler "düşük kopya tekrarları" (İng. low copy repeats ; LCR) olarak adlandırılırlar, çünkü az tekrar eden dizilerdir. Bunlar genelde kromozomun perisentromerik (sentromer çevresi), subtelomerik (telomer yakını) ve interstisyal bölgelerinde bulunurlar. LCR'ler, büyüklükleri (>1Kb), benzerlikleri ve doğrultuları nedeniyle, ikilenmeye ve silinmeye çok müsaittirler. Bu genomik düzen değişikliklere non-alelik homolog rekombinasyon mekanizması neden olur. Bunun sonucu genomik varyasyonlar gen dozajına bağlı hastalık ve bozukluklara yol açabilir, Rett-benzeri sendrom ve Pelizaeus-Merzbacher hastalığı gibi.[7]
Gen duplikasyonu ve amplifikasyonu
Gen duplikasyonu bir canlının genomunda mutlaka uzun vadeli bir değişikliğe yol açmaz. Hatta, bu değişiklikler çoğu zaman ilk meydana geldikleri organizmadan öteye gitmezler. Amplifikasyon bir genin aşırı ifade edilebildiği pek çok yoldan sadece biridir. Doğal bir ikilenme bir somatik hücrede olursa uzun vadeli bir etkisi olmaz, bunun için ikilenmenin bir germ hücresinde meydana gelmesi gerekir. Her durumda, ikilenme kısmen veya ciddi oranda zararlı olabilir. Örneğin, onkogenlerin duplikasyonu çoğu kanser tipinin sık görülen bir nedenidir, göğüs kanserinde P70-S6 Kinaz 1 amplifikasyonunda olduğu gibi.[8] Böyle durumlarda gen düplikasyonu somatik hücrelerde meydana gelir ve sadece kanser hücrelerinin genomunu etkiler, tüm organizma veya onun yavrularının genomu üzerinde etki etmez.
Genomik mikrodizilimlerle ikilenme teşhisi
Genomik mikrodizilim gibi teknolojiler mikroikilenme gibi kromozom anormalliklerini yüksek debili bir yolla teşhis etmekte kullanılır. Bu yöntemde, genom dizisi boyunca yer alan oligonükleotitlerden oluşan bir mikrodizilim çipi üzerinde, test edilecek genomik DNA'nın fluoresan işaretli parçaları hibritlenir. Her bir oligonükleotitteki hibridizsyon sinyali, genomik koordinatlara göre grafiklendiği zaman, normalin altında veya üstünde sinyal gösteren bölgeler, sırasıyla, delesyon veya düplikasyon olan bölgelere karşılık gelir.[9][10]
Kaynakça
- Zhang J (2003). "Evolution by gene duplication: an update". Trends in Ecology & Evolution. 18 (6). ss. 292-8. doi:10.1016/S0169-5347(03)00033-8.
- "Arşivlenmiş kopya". 6 Mart 2014 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Ağustos 2009.
- Taylor JS, Raes J (2004). "Duplication and divergence: the evolution of new genes and old ideas". Annu. Rev. Genet. Cilt 38. ss. 615-43. doi:10.1146/annurev.genet.38.072902.092831. PMID 15568988.
- Ohno, S. (1970). Evolution by gene duplication. Springer-Verlag. ISBN 0-04-575015-7.
- Ohno, S. (1967). Sex Chromosomes and Sex-linked Genes. Springer-Verlag. ISBN 91-554-5776-2.
- Kellis M, Birren BW, Lander ES (Nisan 2004). "Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae". Nature. 428 (6983). ss. 617-24. doi:10.1038/nature02424. PMID 15004568.
- Lee JA, Lupski JR (Ekim 2006). "Genomic rearrangements and gene copy-number alterations as a cause of nervous system disorders". Neuron. 52 (1). ss. 103-21. doi:10.1016/j.neuron.2006.09.027. PMID 17015230.
- "Entrez Gene: RPS6KB1 ribosomal protein S6 kinase, 70kDa, polypeptide 1". 8 Ekim 2009 tarihinde kaynağından arşivlendi.
- Mao R, Pevsner J (2005). "The use of genomic microarrays to study chromosomal abnormalities in mental retardation". Ment Retard Dev Disabil Res Rev. 11 (4). ss. 279-85. doi:10.1002/mrdd.20082. PMID 16240409.
- Gu X, Zhang Z, Huang W (Ocak 2005). "Rapid evolution of expression and regulatory divergences after yeast gene duplication". Proc. Natl. Acad. Sci. U.S.A. 102 (3). ss. 707-12. doi:10.1073/pnas.0409186102. PMC 545572 $2. PMID 15647348.
Ayrıca bakınız
- Psödogenler
- Moleküler evrim
- İnsan genomu
- Karşılaştırmalı genomiks
- Duplikasyon
Dış bağlantılar
- A bibliography on gene and genome duplication31 Ağustos 2012 tarihinde Wayback Machine sitesinde arşivlendi. (Gen ve genom duplikasyonu için bibliyografya)