MikroRNA
Genetikte, mikroRNA (miRNA) yaklaşık 21-23 nükleotit uzunluğunda tek iplikli RNA molekülü türüdür, gen ifadesinin düzenlenmesinde rol oynar. miRNA'lar kodlamayan RNA'lardandır, yani DNA'dan transkripsiyonu yapılan ama proteine çevirisi yapılmayan genler tarafından kodlanırlar. Pri-miRNA olarak adlandırılan primer transkriptler işlenerek, önce pre-miRNA adlı kısa sap-ilmik yapılarına, sonra da fonksiyonel miRNA'ya dönüşürler. Olgun miRNA moleküller bir veya daha çok mesajcı RNA (mRNA) ile kısmî tamamlayıcıdır ve başlıca işlevleri gen ifadesini aşağı ayarlamaktır. 1993'te Lee ve çalışma arkadaşları tarafından Victor Ambros laboratuvarında keşfedilmişlerdir,[1] ancak mikroRNA terimi ilk 2001'de kullanıma girimiştir.[2]
Oluşumu ve işlenmesi
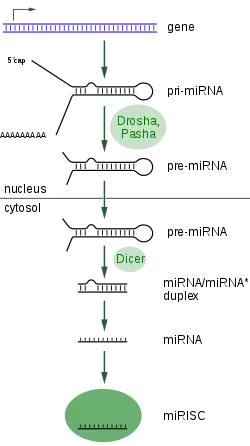
miRNA'yı kodlayan genler işlenmiş olgun miRNA molekülünden çok daha uzundur. miRNA'lar önce birincil (primer) transkript, veya pri-miRNA olarak yazılırlar, bu transkriptlerin birer başlığı ve poli-A kuyruğu vardır. Bunlar işlem görüp hücre çekirdeğinde pre-miRNA olarak bilinen kısa, 70 nükleotit uzunlukta, sap-ilmik şekilli, öncül (prekürsör) yapılara dönüşür. Bu işlenme hayvanlarda Mikroişlemci kompleks (İng. Microprocessor complex) adlı bir protein kompleksi tarafından gerçekleştirilir. Mikroişlemci kompleks'te Drosha adlı bir nükleaz, ve Pasha adlı çift iplikli RNA bağlayıcı protein bulunur.[3]
Pre-miRNA'lar sonra sitoplazmada Dicer adlı endonükleaz ile etkileşerek olgun miRNA'ya dönüşürler. Dicer aynı zamanda RNA-indüklenmiş susturma kompleksi (İng. RNA-induced silencing complex; RISC) oluşumunun başlatır.[4] Bu kompleks miRNA ifadesi ve RNA interferanstan kaynaklanan gen susturmasından sorumludur.
Bitkilerde miRNA oluşumunun yolu, Drosha'nın homologlarının olmamasından dolayı hayvanlardakinden biraz farklıdır; onun yerine Dicer homologları tek başlarına birkaç işlem aşamasını yürütürler.[5] Ayrıca intronik sap-ilmiklerden meydana gelen miRNA'ların oluşumunda da Drosha değil, Dicer görev alır.[6] DNA'nın anlamlı veya ters anlamlı ipliği de miRNA'nın oluşumunda kalıp işlevi görebilir.[7]
Pre-miRNA'nın verimli bir şekilde işlenmesi için, sap-ilmik yapısının hem 5', hem de 3' ucunda tek iplikli RNA uzantıların olması gerekir.[8] Bu tek iplikli RNA motiflerin bileşimlerinden çok uzunlukları son derece önemlidir, işlenmenin gerçekleşebilmesi için. İnsan ve sinek pri-miRNA'larının bir biyoenformatik analizi, çok benzer yapısal bölgelerin varlığını göstermiş, bunlar, bazal kısımlar, aşağı saplar, yukarı saplar ve uç ilmikler olarak adlandırılmıştır; evrimsel olarak korunmuş bu yapılara dayanarak pri-miRNA'nın termodinamik profilleri tanımlanmıştır.[9] Drosha kompleksi, RNA molekülünü uç ilmikten yaklaşık 22 nükleotit uzaktan keser.[10] Çoğu pre-miRNA'da tepesinde bir ilmik olan mükemmel çift sarmallı bir yapı yoktur. Bu seçiciliğin birkaç olası açıklaması vardır. Bir olasılık, 21 baz çiftinden uzun çift sarmallı RNA'nın enterferon tepkisi ve hücrenin anti-viral mekanizmasını harekete geçirmesidir. Bir diğer makul açıklama pre-miRNA'nın termodinamik profilinin hangi ipliğin Dicer kompleksine dahil olacağını belirlemesidir.
Dicer pre-miRNA sap-ilmiğini kestikten sonra iki tamamlayıcı kısa RNA molekülü meydana gelir, ama bunlardan sadece biri RISC kompleksine dahil olur. RISC kompleksinin içinde yer alan bir RNAz olan argonaute'un etkisiyle bu ikisinden 5' ucu daha kararlı olanı seçilip komplekse dahil olur. Bu iplik kılavuz iplik (İng. guide strand) olarak adlandırılır.[11] Öbür iplik, anti-kilavuz veya yolcu iplik olarak adlandırılır, RISC kompleksinin substratı olarak sindirilir.[12] Aktif RISC kompleksine entegre olduktan sonra miRNA'lar kendi tamamlayıcı mRNA molekülleri ile baz eşleşmesi yapar ve argonaute proteinleri tarafından mRNA'nın yıkımına neden olurlar.
Hücresel işlevler
miRNA'nın işlevinin gen düzenlemesi olduğu anlaşılmaktadır. Bu amaçla bir miRNA bir veya daha çok mRNA'yı tamamlayıcıdır (komplemanterdir). Hayvan miRNA'ları genelde 3' UTR bölgesine tamamlayıcıdırlar, bitki miRNA'ları ise mRNA'ların protein kodlayıcı kısımlarına tamamlayıcıdır. miRNA'nın mRNA ile eşleşmesi bazen protein çevirisini engeller ve bazen de mRNA'nın kesilmesini (RNA interferansa benzer bir süreçle) kolaylaştırır. Çoğu hayvan miRNA'sı protein çevirisini engeller, çoğu bitki miRNA'sı da mRNA'yı keserek çalışır. miRNA'lar ayrıca hedef mRNA'ya karşılık gelen genomik bölgelerde DNA metilasyonuna neden olabilirler. miRNA'lar kendilerini tamamlayan bir grup proteinle (mikroribonükleoproteinler = miRNP) birlikte işlev görürler.
miRNA'nın etkileri ilk 1993'te Victor Ambros ve çalışma arkadaşları tarafından C. elegans solucanında keşfedildi.[1] miRNA'ların varlığı çeşitli bitki ve hayvanlarda teyid edilmiştir. Ökaryotik miRNA genlerinin benzerleri bakterilerde de bulunmuştur, bunlar mRNA ile eşleşerek mRNA çokluğunu ve çevirisini kontrol etmektedirler, ancak bu süreçte Dicer enziminin bir benzeri yer almadığı için bu RNA'lar genel olarak miRNA olarak sayılmamaktadırlar.
Bitkilerde kısa müdaheleci RNA'lar (İng. short-interfering RNA; siRNA kısaltmasıyla anılır) viral RNA'nın transkripsiyonunu engellemeye yarar. siRNA çift iplikli olmasına rağmen, etki mekanizması miRNA'nınkine yakından benzer, saç firkete yapıları göz önüne alınırsa bu benzerlik daha da çarpıcıdır. siRNA'lar, miRNA'lar gibi, gen denetimine yararlar.
Gen aktivasyonu
Çift iplikli RNA (İng. double stranded RNA'dan dsRNA olarak kısaltılır) gen ifadesini ayrıca etkinleştirebilir de, buna "küçük RNA indüklenmiş gen aktivasyonu" (İngilizce small RNA-induced gene activation veya RNAa). Gen promotörlerinin dsRNA ile hedeflenmesi, ilişkili genlerde güçlü bir transkripsiyon aktivasyonu yaratabilir. İnsan hücrelerinde bu hem sentetik dsRNA ile ("küçük etkinleyici RNA" veya İng. small activating RNA veya saRNA),[13] hem de endojen miRNA için gösterilmiştir.[14]
mRNA sinyalizasyonunun belirlenmesi ve manipülasyonu
Bir miRNA'nın aktivitesi bir kilitli nükleik asit türü olan bir morfolino veya 2'-O-metil RNA oligo kullanılarak deneysel olarak bloke edilebilir.[15][16][17] miRNA'nın olgunlaşmasındaki adımlar, bloke edici oligonükleotitler ile engellenebilir.[18] Bir miRNA'nın mRNA üzerindeki bağlanma yeri de bir oligo tarafından sterik olarak bloke edilebilir.[19][20].
miRNA ve hastalık
miRNA ökaryotik hücrelerin normal işlevinde yer aldığı gibi, miRNA'nın bozuk çalışması da hastalığa neden olur. Kanser, nöroloji, kardiyoloji, viroloji'de bu konuda çalışmalar sürmektedir.
miRNA ve kanser
Birkaç miRNA ile bazı kanser tipleri arasında ilişkiler bulunmuştur.
Aşırı c-myc (çeşitli kanserlerde rol oynayan bir protein) üreten bir fare türünde yapılan araştırmalar miRNA'nın kanser gelişmesine etki ettiğini göstermiştir. Lenfomalarda bulunan miRNA'ları özellikle çok miktarda üretmek üzere tasarlanmış farelerde 50 gün içinde kanser gelişmiş ve bunlar iki hafta ardından ölmüşlerdir.[21]
Bir diğer çalışmada, hücre çoğalmasını düzenleyen E2F1 proteininin iki tip miRNA tarafından inhibe edildiği gösterilmiştir. miRNA, mesajcı RNA'ya bağlanarak gen aktivitesine etki eden proteinlerin çevrimini engellemektedir.[22]
miRNA kodlayan 217 genin etkinliği ölçülerek farklı kanser tiplerini ayırdebilen gen aktivite örüntüleri bulunmuştur. miRNA profilleri kanserlerin sınıflandırılmasına yarayabilir. Böylece hekimler bir kanser dokusunu analzi edip onun hangi dokudan yayılmış olduğunu anlayabilir ve özgün doku tipine göre tedavi planlayabilirler. Kronik lenfositik lösemili hastaların bu kanserin hızlı mı yavaş mı gelişen türünü taşıdıkları anlamak için miRNA profillemesi kullanılabileceği gösterilmiştir.[23]
miRNA ve kalp hastalığı
miRNA fonksiyonunun kalpteki global rolünün anlaşılması için fare kalbinde miRNA olgulaşması engellenmiş ve miRNA'nın bu organın gelişmesinde önemli bir rol oynadığı görülmüştür.[24][25] miRNA expression profiling studies demonstrate that expression levels of specific miRNAs change in diseased human hearts, pointing to their involvement in cardiomyopathies.[26][27][28] Ayrıca, belli miRNA'lar üzerinde yapılan araştırmalar kalp gelişimi sırasında miRNA'ların farklı rolleri olduğunu göstermiştir.[25][29][30][31][32][33]
Kaynakça
- Lee RC, Feinbaum RL, Ambros V (1993). "lin-4". Cell. Cilt 75. ss. 843854. doi:10.1016/0092-8674(93)90529-Y. PMID 8252621. 4. harf sırasında bulunan
|sayfalar=parametresi C1 control character içeriyor (yardım) - Ruvkun, G. (Oct 26 2001). "Molecular biology. Glimpses of a tiny RNA world". Science. 294 (5543). ss. 7979. doi:10.1126/science.1066315. PMID 11679654. 2 Haziran 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Eylül 2008. 4. harf sırasında bulunan
|sayfalar=parametresi C1 control character içeriyor (yardım); Tarih değerini gözden geçirin:|yıl=(yardım) - Denli AM, Tops BB, Plasterk RH, Ketting RF, Hannon GJ. (2004). Nature 432(7014):231-5.
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ. (2001). Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409(6818):363-6.
- Kurihara Y, Watanabe Y. (2004). Arabidopsis micro-RNA biogenesis through Dicer-like 1 protein functions. Proc Natl Acad Sci USA 101(34):12753-8.
- Gao F-B (2007). "Posttranscriptional control of neuronal development by microRNA networks". Trends in Neurosciences. Cilt 31. s. 20. doi:10.1016/j.tins.2007.10.004.
- Stark A, Bushati N, Jan CH; ve diğerleri. (2008). "A single Hox locus in Drosophila produces functional microRNAs from opposite DNA strands". Genes Dev. 22 (1). ss. 813. doi:10.1101/gad.1613108. PMID 18172160. 2. harf sırasında bulunan
|sayfalar=parametresi C1 control character içeriyor (yardım) - Zeng Y, Cullen BR (2005). "Efficient processing of primary microRNA hairpins by Drosha requires flanking nonstructured RNA sequences". J. Biol. Chem. 280 (30). ss. 27595603. doi:10.1074/jbc.M504714200. PMID 15932881. 6. harf sırasında bulunan
|sayfalar=parametresi C1 control character içeriyor (yardım) - Han J, Lee Y, Yeom K-H, Nam J-W, Heo I, Rhee J-K, Sohn SY, Cho Y, Zhang B-T, Kim VN (2006). "Molecular basis for the recognition of primary microRNAs by the Drosha-DGCR8 complex". Cell. 125 (5). ss. 887901. doi:10.1016/j.cell.2006.03.043. 4. harf sırasında bulunan
|sayfalar=parametresi C1 control character içeriyor (yardım) - Zeng Y, Yi R, Cullen BR (Ocak 2005). "Recognition and cleavage of primary microRNA precursors by the nuclear processing enzyme Drosha". EMBO J. 24 (1). ss. 13848. doi:10.1038/sj.emboj.7600491. PMC 544904 $2. PMID 15565168. 4. harf sırasında bulunan
|sayfalar=parametresi C1 control character içeriyor (yardım) - Preall JB, He Z, Gorra JM, Sontheimer EJ. (2006). Short interfering RNA strand selection is independent of dsRNA processing polarity during RNAi in Drosophila. Curr Biol 16(5):530-5.
- Gregory RI, Chendrimada TP, Cooch N, Shiekhattar R. (2005). Human RISC couples microRNA biogenesis and posttranscriptional gene silencing. Cell 123(4):631-40.
- Li LC (2008). "Small RNA-Mediated Gene Activation". RNA and the Regulation of Gene Expression: A Hidden Layer of Complexity. Caister Academic Press. ISBN 978-1-904455-25-7.
- Place RF, Li LC, Pookot D, Noonan EJ, Dahiya R (2008). "MicroRNA-373 induces expression of genes with complementary promoter sequences". Proc. Natl. Acad. Sci. U.S.A. 105 (5). ss. 160813. doi:10.1073/pnas.0707594105. PMID 18227514. 5. harf sırasında bulunan
|sayfalar=parametresi C1 control character içeriyor (yardım) - Kloosterman, WP (Dec 7 2004). "Substrate requirements for let-7 function in the developing zebrafish embryo". Nucleic Acids Res. 32 (21). ss. 628491. doi:10.1093/nar/gkh968. PMID 15585662. 5. harf sırasında bulunan
|sayfalar=parametresi C1 control character içeriyor (yardım); Tarih değerini gözden geçirin:|yıl=(yardım) - Flynt, AS (2007). "Zebrafish miR-214 modulates Hedgehog signaling to specify muscle cell fate". Nature Genetics. Cilt 39. ss. 259263. doi:10.1038/ng1953. PMID 17220889. 4. harf sırasında bulunan
|sayfalar=parametresi C1 control character içeriyor (yardım) - Meister, G (Mart 2004). "Sequence-specific inhibition of microRNA- and siRNA-induced RNA silencing". RNA. 10 (3). ss. 54450. doi:10.1261/rna.5235104. PMID 14970398. 4. harf sırasında bulunan
|sayfalar=parametresi C1 control character içeriyor (yardım) - Kloosterman, WP (2007). "Targeted inhibition of miRNA maturation with morpholinos reveals a role for miR-375 in pancreatic islet development". PLoS Biol. (Pubmed). 5 (8). ss. e203. doi:10.1371/journal.pbio.0050203. PMID 17676975.
- Choi, WY (2007). "Target Protectors Reveal Dampening and Balancing of Nodal Agonist and Antagonist by miR-430" (Pubmed). Science. PMID 17761850.
- Klein, ME (2007). "Homeostatic regulation of MeCP2 expression by a CREB-induced microRNA" (– Scholar search). Nature Neuroscience. Cilt 10. s. 1513. doi:10.1038/nn2010.
- He L, Thomson JM, Hemann MT, Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe SW, Hannon GJ, Hammond SM (2005). "A microRNA polycistron as a potential human oncogene". Nature. 435 (7043). ss. 828-833. doi:10.1038/nature03552. PMID 15944707.
- O'Donnell KA, Wentzel EA, Zeller KI, Dang CV, Mendell JT (2005). "c-Myc-regulated microRNAs modulate E2F1 expression". Nature. 435 (7043). ss. 839-843. doi:10.1038/nature03677. PMID 15944709.
- Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA, Downing JR, Jacks T, Horvitz HR, Golub TR (2005). "MicroRNA expression profiles classify human cancers". Nature. 435 (7043). ss. 834-838. doi:10.1038/nature03702. PMID 15944708.
- Chen JF, Murchison EP, Tang R; ve diğerleri. (Şubat 2008). "Targeted deletion of Dicer in the heart leads to dilated cardiomyopathy and heart failure". Proc. Natl. Acad. Sci. U.S.A. 105 (6). ss. 2111-2116. doi:10.1073/pnas.0710228105. PMID 18256189.
- Zhao Y, Ransom JF, Li A; ve diğerleri. (Nisan 2007). "Dysregulation of cardiogenesis, cardiac conduction, and cell cycle in mice lacking miRNA-1-2". Cell. 129 (2). ss. 303-317. doi:10.1016/j.cell.2007.03.030. PMID 17397913.
- Thum T, Galuppo P, Wolf C; ve diğerleri. (Temmuz 2007). "MicroRNAs in the human heart: a clue to fetal gene reprogramming in heart failure". Circulation. 116 (3). ss. 258-67. doi:10.1161/CIRCULATIONAHA.107.687947. PMID 17606841.
- van Rooij E, Sutherland LB, Liu N; ve diğerleri. (Kasım 2006). "A signature pattern of stress-responsive microRNAs that can evoke cardiac hypertrophy and heart failure". Proc. Natl. Acad. Sci. U.S.A. 103 (48). ss. 18255-60. doi:10.1073/pnas.0608791103. PMID 17108080.
- Tatsuguchi M, Seok HY, Callis TE; ve diğerleri. (Haziran 2007). "Expression of microRNAs is dynamically regulated during cardiomyocyte hypertrophy". J. Mol. Cell. Cardiol. 42 (6). ss. 1137-41. doi:10.1016/j.yjmcc.2007.04.004. PMID 17498736.
- Zhao Y, Samal E, Srivastava D (Temmuz 2005). "Serum response factor regulates a muscle-specific microRNA that targets Hand2 during cardiogenesis". Nature. 436 (7048). ss. 214-20. doi:10.1038/nature03817. PMID 15951802.
- Xiao J, Luo X, Lin H; ve diğerleri. (Nisan 2007). "MicroRNA miR-133 represses HERG K+ channel expression contributing to QT prolongation in diabetic hearts". J. Biol. Chem. 282 (17). ss. 12363-7. doi:10.1074/jbc.C700015200. PMID 17344217.
- Yang B, Lin H, Xiao J; ve diğerleri. (Nisan 2007). "The muscle-specific microRNA miR-1 regulates cardiac arrhythmogenic potential by targeting GJA1 and KCNJ2". Nat. Med. 13 (4). ss. 486-91. doi:10.1038/nm1569. PMID 17401374.
- Carè A, Catalucci D, Felicetti F; ve diğerleri. (Mayıs 2007). "MicroRNA-133 controls cardiac hypertrophy". Nat. Med. 13 (5). ss. 613-8. doi:10.1038/nm1582. PMID 17468766.
- van Rooij E, Sutherland LB, Qi X, Richardson JA, Hill J, Olson EN (Nisan 2007). "Control of stress-dependent cardiac growth and gene expression by a microRNA". Science (journal). 316 (5824). ss. 575-9. doi:10.1126/science.1139089. PMID 17379774.
Ayrıca bakınız
- Gen ifadesi
- RNAi
- siRNA